- UID
- 256426
- 精华
- 1
- 帖子
- 217
- 最后登录
- 2015-3-25
- 注册时间
- 2012-5-30

海 水
童 生

铭
鱼



 龙币 龙币- 21179
 兑换福利 兑换福利
|
本帖最后由 有鲤有我 于 2013-7-14 14:55 编辑
这篇文章堪称经典!介绍了海水白点的成因,及多种有效的治疗方法。本篇為全文五部曲中的第一部。第二部将详细说明海水白点虫的生命周期、如何辨别是否感染、表列出临床徵状、其适应能力,以及面对白点虫的新挑战及预防方式。(鱼病治疗) 此篇作者:lammark
第一章

白点病治疗
引言
海水白点虫(Cryptocaryon irritans)向来被认定為是侵袭海水鱼类中,破坏力最强的寄生虫之一(Wright & Colorni, 2002),其高感染力长期来造成不论是观赏鱼类或食用鱼类的死亡以及饲主的经济损失。尤其在水族箱中因系一封闭环境,更是提供了白点虫一个绝佳的繁殖环境。在一封闭系统中,预估白点虫每6至8天,其数量便可繁殖近10倍之多(Burgess, 1992)。
尽管我们对於此一寄生虫已有相当了解,事实上白点虫目前仍是对於饲养硬骨海水鱼类健康方面的最大威胁之一。此原因有部份便是由於许多海水鱼玩家及销售业者,长期以来对於白点虫的生命周期、传染模式及治疗方式所传递的资讯,事实上并不正确(虽然不是有意的)。本文共分為五篇,目的便在详述白点虫的各方面资讯,希望能藉此导正那些原本错误的认知。
什麼是海水白点虫(Cryptocaryon irritans)?
Cryptocaryon irritans一般被海水鱼业者称為“ICH”或是“白点”,它是一种发现在热带与温带海洋硬骨鱼类身上,附有纤毛的原生寄生虫。此病原体系海水鱼类之专性寄生虫(Iwama, Pickering, Sumpter & Shreck, 1997. Dickerson & Dawe, 1994),亦即因其系依靠鱼类取得食物来源,必须依附在鱼类身上才能成长,因此在其寄生阶段(theront stage)若是无法依附於宿主,此寄生虫将只能存活一至二日。
歷史
1937年,日本人Sikama首次提出关於海水白点虫的报告,起因於这中被称為纤毛寄生体的白点虫,感染了东京帝国大学水產学会水族箱中的45种鱼类。在此之前,此一寄生虫的感染行為仅止於在公立或私人的水族馆中观察(Nigrelli & Ruggieri, 1966. Wilkie & Gordin, 1969)。自此以后,海水白点虫的问题,便开始在海洋生物养殖上逐渐受到重视(Colorni, 1985. Huff & Burns, 1981)。海水白点虫与淡水白点虫有许多相近之处,但事实上他们是属於两种完全不同分类的寄生虫。
近期发展
广泛的系统分析以及分子诊\断方法被使用於海水白点虫的重新归类。近来此一原生体外寄生虫被建议应归入Prostomatea「纲」中的Prorodontida 「目」,且另新增一「科」Cryptocaryonidae做為归类(Wright & Colorni, 2002)。
研究团体近来发现一些令人担忧的新证据,显示出海水白点虫具有适应新环境的能力。他们尝试将白点虫置於数种不具有生存压迫的环境中,迫使白点虫必须不断的适应新环境及变化(Yambot, et al., 2003)。实验结果却因而產生数个新的品种出来(Diggles & Adlard, 1995, 1997 Diggles & Lester, 1996a,b,c. Jee et al., 2000)。同时白点虫也在一些新的地区被发现,这些地区的水温仅有12~16℃,较先前报告中的温度為低(Jee, et al., 2000)。在过去,白点虫被认為无法生存於19℃以下的环境中(Nigrelli & Ruggieri, 1966. Diggles & Lester, 1996b. Gordin, 1969)。而在臺湾属於低盐的环境中,也发现了高度变异的品种已在大量繁殖(Yambot et al., 2003)。由上述新发现我们得知,海水白点虫事实上在不同地理区域、不同水温及不同盐度中都能生存,因此我们必须要有新的策略的疗法来控制它。
迷思与误解
海水玩家普遍存有相同的误解,以為只有健康状况较差的鱼只才容易被白点虫感染。当然,在压迫的环境下,鱼只的免疫功能会降低,使得鱼只在受到感染后,无法產生所需的免疫力。然而压迫的环境或是鱼只健康情形不佳,并非是否会感染白点病的必要因素,即使是健康的鱼只在未受压迫的情形下,若是暴露在白点虫的环境中,仍然狠容易受到感染。值得注意的是,在临床实验中,「紧迫」并非鱼只是否会感染白点病的因素之一。
另一个在海水玩家中普遍存在的误解是认為白点虫在每一个鱼缸中或多或少一定会有,其实他们是误将白点虫视為与缸中普遍存在的细菌或病原体同类,因而假设这些原本已存在的白点虫,在缸子状况不好时(如水质不好或是喂食污染)便会爆发在鱼只身上。抱持这种看法的人通常都忽视了新鱼只在入缸前检疫的重要性,白点病必须由外来鱼只引进,否则无法在鱼缸中发生。一般说来,白点病是藉由已被感染的鱼只所带入的,未针对新进鱼只进行检疫,将大大提高传染的可能性。而由於缸中一般还有活石或软体生物,一旦白点病爆发后将更难处理。
Dr. Harry W. Dickerson在1994年夏季版的Seascope杂志中,针对海水白点虫做了以下论述:「要挽救缸中生物,实际上必须完全消灭所有的寄生虫,治疗才能產生效果」(Dickerson, 1994)「根除海水缸或养殖场白点虫的难题来自於其生命周期的復杂性,特别是有些白点虫在分裂体 Tomonts以及非同步脱离孢囊具传染性的掠食体阶段的潜伏期极长。」(Colorni & Burgess, 1997)但透过有效的治疗与适当的程序,白点病仍可从鱼缸及鱼只身上完全消除。
.
海水白点虫属於专性寄身虫的一种,亦即在找不到宿主的情形下,它便无法完成其生命周期(在营养体阶段)。考虑到白点虫在分裂体Tomonts阶段需要3到28天孵化,因此建议将鱼只清空30天到6周的时间,使缸中找不到宿主,便能将白点虫从缸中完全消灭。并将移至治疗缸的鱼只施以有效的治疗(如使用低比重疗法或下铜药),以消灭鱼只上的白点,可将其治癒。
哪些鱼容易被感染?
海水白点虫并无特定的宿主选择性(Burgess & Mathews, 1995),这表示了只要暴露在有白点虫的环境中,任何种类的鱼只都可能会被感染。甚至原本并非海水鱼类亦同,海水化的黑茉莉鱼就曾被用来做為试验品,以了解海水白点虫是否需要特定宿主(Yoshinaga & Dickerson, 1994)。此外,在鱼缸或封闭系统中,白点虫繁殖的速度将以倍数激增,使得要对抗它愈来愈困难。鱼缸中之所以较大海中容易传染白点病,系由於鱼缸中的相对高密度养殖,使得白点虫在掠食体Theronts阶段容易找到宿主(Yambot, et al., 2003)。
狠明显的,不同种的礁巖鱼类对於白点病有不同程度的抵抗力(Colorni & Burgess, 1997. Diggles & Lester, 1996c),在封闭的环境中,其抵抗力因素可能远比不同鱼种对食物或环境需求等种种因素要来得重要。一般来说,神仙类、倒吊类、鰈类或箱魨科鱼类,通常被认為较容易感染白点病,而板鳃类例如鯊鱼或是魟鱼等,则天生对白点病具有抵抗力(Lom, 1984)。
传染模式
海水白点病通常起因於鱼缸中所加入的新鱼只原本就已经受到感染(Dickerson & Dawe, 1995),已受感染的鱼只通常在身体上会发现有白点或是伤口,也可能因為病原体仍在营养体Trophonts阶段,宿於鳃膜中而无法从外观察觉。
虽然并不常见,但白点病也可能是藉由其他管道进入鱼缸中,例如所置换的海水中存有正处於游离阶段的白点虫掠食体(Theront)。然而,由於孵化出来的掠食体Theront能单独存活的时间极短,因此藉由这个管道导致感染的机会其实并不高,只要将海水静置24小时后再使用,基本上便可免除这个可能性(Colorni & Burgess, 1997)。任何表面坚硬的物体,如砂子、石头、玻璃、缸中设备,甚至无脊椎生物(虽然他们不会感染白点病),都可让白点虫在分裂体Tomonts阶段做為着床物(Burgess, 1992),所以假若你将这些物体移到另一个乾凈的缸中,则分裂体Tomonts或囊胞也会有可能被带入。同时,使用海水活体喂食时,理论上也可能因此带入白点虫。
风乾鱼缸、砂子、石头与设备可以除去附着在上面的分裂体Tomonts,但狠显然的,这个方法并不适用在活石上。倘若取得软体、活石或活砂的来源处已被感染白点病,那麼最好还是先经过检疫的程序后再移入缸中。
致死的原因
在一重度感染的封闭环境中,海水白点虫可引发鱼只的大量死亡(Yoshinaga & Dickerson, 1994)。海水白点虫会引发之主要病变,称為白点病;其次还有某种细菌或真菌的感染,通常也与白点虫有关。海水白点虫会导致宿主因窒息、渗透不平衡(指细胞膜内外液体压力不平衡)以及继发性细菌感染而死亡。
白点病会使得鱼只鳃膜產生上皮细胞肥大的徵状以及细胞次胶层的融化(Yambot, et al., 2003),若是攻击鱼只的白点虫数量足够,在受到感染的初期,肉眼可视鱼只有白点產生,而鱼鳃上皮将会严重受损(Dickerson & Dawe, 1994);若是病状严重或遭受重復感染,将严重导致细胞次胶层融化及各薄膜间空间不可逆的闭合 (Colorni & Burgess, 1997)。受掠食体(Theronts)侵袭所造成的伤口经常会成為再次為伺机性细菌或是真菌病原体感染的途径(Colorni & Burgess, 1997),这些伤口也特别容易為假单胞类细菌感染而引发并发癥(Nigrelli & Ruggieri, 1966)。同时这些伤口也会危急鱼只黏液/鳞片/表皮间的平衡,使得渗压功能无法正常作用及更為耗费鱼只的体力,对严重感染的鱼只而言,更可能造成渗透压的剧烈改变。一般说来,鱼只仅在遭受连续性严重感染后,才会导致死亡(Colorni, 1992)。
|
|
第二章
本篇是為关於海水型白点虫 (亦称為 “盐水型白点虫saltwater ich.”)的五篇系列文章中的第二章. 第一章的部份包含了关於海水型白点虫的介绍, 以及其歷史, 还有相关的研究发展. 也介绍了许多对於白点虫的迷思及误解: 什麼样的鱼种易受感染, 感染模式及致死的原因. 本章将会继续接着介绍海水型白点虫的生命周期, 如何辨认海水型白点虫, 感染的临床癥状, 适应变异的能力, 新的挑战及预防的方法
生命周期
海水型白点虫的生命周期可区分為明确的四个型态(Colorni & Burgess, 1997). 海水型白点虫并不像其他鱼类寄生虫般的需要中间宿主(例如: 蜗牛等). 四个型态的生活周期主要可以区分為寄生期及脱离宿主后两大阶段. 这四个型态是為掠食体 theront, 原分裂前体protomont,分裂体 tomont 及 营养体trophont. 海水型白点虫的生命周期在温度為24-27C 的情况下, 通常大约是在1到2周左右(Colorni, 1992). 生命周期的长度会因為变异种或特有种而有些微的差异(Colorni & Burgess, 1997). 到至今有纪录的文件中并未发现在其生命周期中有任何休眠状态的存在. 然而, 海水型白点虫分裂体tomonts 所需要的孵化时间约為 3 到 28 天(Colorni, 1985). 在纪录中 分裂体tomonts 孵化所需的最长时间為 72 天(Colorni & Burgess, 1997). 然而海水
型白点虫的生命周期与温度是有相关联的, 所以这样长的孵化时间并不可能存在於热带鱼缸中的.
白点病治疗
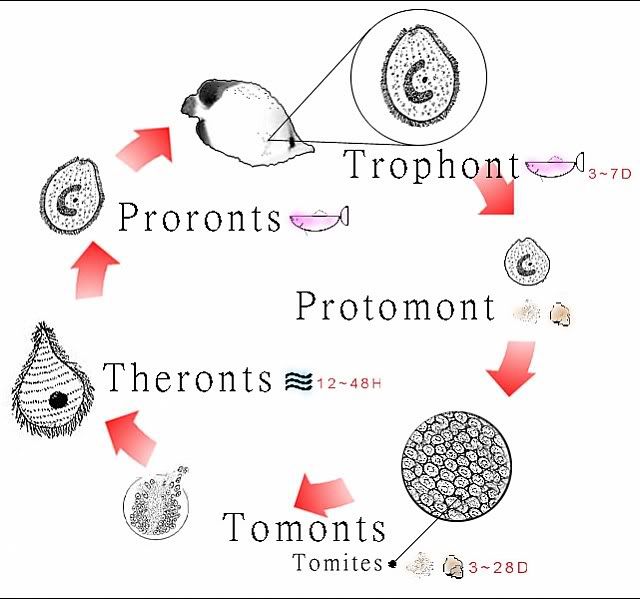
PS.这是热心的Brick版主提供自己精心制作的生命史图解图片
通常当鱼身上被用来当為癥状指标的白点短暂退去的时候, 鱼友常误以為白点感染已经治癒. 事实上, 在白点虫的生命周期里, 当 营养体Trophonts 成熟后, 便会离开宿主进入所谓分裂体 tomonts 繁殖的阶段. 一般来说, 白点的消失只是短暂的, 通常几天过后, 会有更多白点出现.
.
营养体Trophont 或寄生摄食阶段
鱼友通常最熟悉的便是海水型白点虫的营养体 trophont 型态. 白点虫的摄食型态乃是以类似盐粒般的白点出现在鱼的身上. 营养体Trophonts 会持续性的依附在宿主身上的体表或上皮组织中. 并摄食宿主鱼身上的体液, 组织碎片, 或体表细胞(Colorni & Burgess, 1997). 并靠着从宿主身上摄食来的营养成长, 逐渐成為以肉眼便可辨识的白点. 通常会在 3 到 7 天里完全成熟并离开宿主, 平均时间多為 4 到 5 天内(Colorni & Diamant, 1993. Colorni, 1985). 在同一宿主鱼身上的成熟的营养体 trophonts 通常会在 16-18小时 的短时间内离开宿主. 如果宿主鱼死亡, 营养体trophonts 也会离开宿主鱼(Dickerson & Dawe, 1994) 若因宿主死亡而被迫在成熟前便离开宿主的营养体 trophonts, 虽然也能够產生分裂体 tomonts, 但这些分裂体 tomonts 常常因為不够成熟而无法產生可存活的掠食体theronts.
.
原分裂前体Protomont 或离开宿主后且未形成胞囊前
营养体Trophonts 在成熟后便会离开宿主, 并退去身上的纤毛. 这就是所谓的原分裂前体 protomont 阶段. 原分裂前体Protomonts 通常会於清晨五点左右或清晨前夕离开宿主鱼(Burgess & Mathews, 1994b). 至於营养体Trophonts 是否利用清晨前的短暂黑暗时间离开宿主, 以减低被攻击风险的生存策略, 在学说理论上尚未被确立. 这样的时间点狠可能只是巧合. 脱离宿主后的 2 到 8 小时,原分裂前体 Protomonts 会在底沙或活石上移动, 直到黏附在适当的地方.
分裂体Tomont 或繁殖阶段
原分裂前体Protomonts 会先吸附在鱼缸中的底砂, 活石, 或其他适合的硬质表面上, 然后开始形成胞囊状的分裂体 tomonts(Colorni, 1985). 这是所谓的繁殖阶段. 在接下来的 8 到 12 小时, 分裂体Tomonts 会在体内开始形成一些子细胞, 也就是仔体 tomites. 每一个分裂体 tomont 所能產生的仔体 tomites 数量, 会因品种的不同而有所差异, 某些品种所能產生的 仔体 tomites 可能小於 200 个, 而有些品种则可能多达上千个(Diggles & Adlard, 1997). 当孕育成熟后, 分裂体tomonts 会开始孵化并将仔体 tomites 释放到海水中. 这时被孵化出来的都是具备游泳能力的单一个体, 称之為掠食体 theronts.
分裂体Tomonts 孵化所需的时间会因某些因素的影响而產生极大的差异, 可能从最短的 3 天到最长的72 天左右(Noga, 2000). 海水型白点虫的生命周期与温度有狠大的相关性, 所以像 72 天这样长的孵化时间基本上说来是非常不寻常的, 且只可能发生在温度较低的环境中. 在适合珊瑚生长的温度下,分裂体 tomonts 孵化的时间约為 3 到 28 天, 平均高峰集中在 4 到 8 天(Colorni, 1985). 这些差异的存在或许是海水型白点虫所演化出来的生存策略. 然而, 在 tomont 型态下两星期后,分裂体 tomont 所能產生的t掠食体heronts 不仅在数量上明显的减少, 且孵化出来的 掠食体theronts 在感染能力上也大幅的衰减(Colorni, 1992).
Theront 掠食体或具感染性的浮游阶段
在海水型白点虫生命周期中具传染性的浮游阶段称之為掠食体 theront. 在不同的研究报告中指出, 从分裂体 tomont 孵化出来后的 12 到 48 小时间, 掠食体Theronts 是存活在水中的(Burgess & Matthews, 1994a.Yoshinaga & Dickerson, 1994. Colorni, 1985). 掠食体Theronts 必须在这段时间内找到适合的宿主, 否则便会死亡, 温度, 比重(盐度), 或特有种的差异可以解释寻求宿主时间上的差异.
掠食体Theronts 孵化的时间, 一致性的都维持在清晨两点到九点之间(Yoshinaga & Dickerson, 1994). 从分裂体 tomont 阶段到掠食体theront 阶段的生理变化周期与光线虽然并没有关系, 但这也是尚未被解释说明的. 在从 分裂体tomont 阶段孵化之后, 掠食体Theronts 便快速的失去感染的能力. 6 到 8 小时候后的感染能力便变的狠低(Burgess, 1992). 孵化后的 7.5 小时, 87%的掠食体 theronts 仍然存活. 经过11.5 小时后, 存活率便只剩下 9%. 孵化后的15.5 小时, 更只剩下 0.34%. (Yoshinaga & Dickerson, 1994). 掠食体Theronts 的大小会因為白点虫品种的不同, 地理上的分布, 宿主的品种差异, 以及水温的变异而有所不同(Colorni & Burgess, 1997). 在 掠食体theront 接触到宿主之后, 开始附着的阶段称之為原营养前体proront.原营养前体 Proronts 在附着之后会开始鉆进宿主鱼的上皮组织, 这些伤口在短时间(可能只要五分鐘)内便会快速的恢復, 并覆盖住原营养前体 Proronts (Colorni & Burgess, 1997). 然后 原营养前体Proronts 便快速的转化成营养体 trophonts, 展开寄生摄食的阶段.
唯一能够确认感染海水型白点虫的方法是用显微镜来观察新鲜鱼鳃, 鱼鰭的切片或从皮肤取得的抹片上, 是否有持续旋转动作, 梨形的纤毛虫(营养体trophonts)的存在(Colorni & Burgess, 1997). 然而对鱼友而言, 如果对於自己所饲养鱼只的外表及日常行為有深切的了解时, 便能藉由观察一些容易辨识的临床癥状来确认是否感染白点虫.
- 可能的临床癥状列表
- 出现针头或句点般大小的白点, 一开始会出现在皮肤或鱼鰭上, 然后出现在眼睛的部位
-通常一些鱼直到疾病已经发展才会显露出受到疾病的侵袭,但不是全部
- 分泌黏液增加
- 在感染初期, 会有过动的行為
- 在鱼缸中有磨擦抓痒, 冲撞的行為
- 有震颤, 抽搐, 痉挛的状况
- 寻找遮蔽物, 或躲藏的行為
- 出现的白点会在消失数天后再次出现
- 除感染早期之外, 会出现呼吸急促的癥状
- 蒙眼癥状產生并伴随着细菌感染
- 鱼体出现退色的情况
- 在感染后期, 出现烂鰭或其他细菌感染癥状
- 皮肤出现因细菌感染所引起溃烂脱色的情况
- 停留在靠近水面或水流强劲的区域
- 缺乏食慾
- 感染后期, 出现脱水及体重急速下降的癥状
海水型白点虫的适应能力
在经过几个世代演化后的分裂体 tomonts 身上, 发现了一些成熟个体特徵的变异. 这些变异的特徵包含分裂体 tomonts 从单一生存的个体, 演进成群聚化.分裂体 Tomonts 也从没有或微弱黏着性演变成具有黏着性 (Yambot, et al., 2003). 依据报导指出, 有一种海水型白点虫冷水变异种, 会藉由出芽繁殖来產生子代的分裂体 tomonts (Jee, et al., 2000). 另外, 在上皮组织中的微小管道中也发现了具有微弱黏着性的 tomonts (Diamant, et al., 1991).
在臺湾及其他地区, 已经确认数种新品系的海水型白点虫的存在(Burgess & Mathews, 1995. Diggles and Adlard,1997).
其中在嘉义及高雄所发现具有高度变异性的特殊种特别的引人注目. (Yambot, et al., 2003) 嘉义特有种是在一个比重(盐度)只有 5ppt 的小池塘中所发现的. 这是在有纪录的文件中, 第一次在这样低的比重(盐度)中发现海水型白点虫. 高雄特有种则是从一个比重(盐度)為10 ppt 的洞穴中所发现的分裂体 tomonts 的第 12 个世代所取得的(Yambot, et al., 2003). Diggles and Lester, (1996a) 提出海水型白点虫的分布范围已经扩散分布到淡水出海口的区域.
新的挑战
海水型白点虫在地理, 温度以及盐度范围上的分布已经令人忧心的愈来愈广大, 且臺湾特有种也相对的显现出海水型白点虫在物种上的多样性演化(Diggles & Adlard, 1997. Yambot, et al., 2003) 这些报告让我们了解到, 实际上海水型白点虫是有足够的适应能力来面对并适应新环境的变异. 更让我们了解到要能有效的控制海水型白点虫病, 新的策略及治疗方式是必要的. 就以往的文献海水型白点虫 各品系 在低比重(盐度)的治疗方式下, 能够有效的消灭(Colorni, 1987. Rigos et al., 2001).
预防措施
相对於治疗被感染的鱼只而言, 事前的预防措施是為较有效及较佳的处理方式. 对所有新进鱼只进行為期至少叁周的隔离检疫, 将能大幅度的预防主缸中海水型白点虫病的爆发. 六周以上的隔离检疫期对於主缸则有更佳的安全保障. 严格的预防措施以及适当的检疫步骤是维持鱼缸免於海水型白点虫病的感染最佳方法(Colorni & Burgess, 1997).
.
如果鱼只能经由隔离检疫的过程来观察其是否有染病的情况, 不仅能让染病鱼只的治疗更形简单, 也相对降低了主缸中已完成检疫治疗鱼群的染病风险. 在初期检疫的过程中, 可以对鱼只施与低比重(盐度)的治疗方式. 这样的处理将巨幅的降低将寄生虫导入主缸的风险. 当主缸中的鱼只因感染海水型白点虫而需被移出主缸进行检疫治疗时, 主缸应采取鱼只凈空的动作(主缸内所有的鱼只都须被移出, 无脊椎动物除外), 為期至少 30 天. 这样的作法是為了要让主缸中的寄生虫因為没有宿主的存在而无法存活. 一般说来, 30 天的主缸凈空期已是足够, 然而, 更长的凈空时间会有更佳的效果, 也相对增加主缸的安全性.
|
|

















 海水鱼白点病症状
海水鱼白点病症状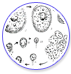 海水鱼白点病原因
海水鱼白点病原因 海水鱼白点病的预防
海水鱼白点病的预防 海水鱼白点病治疗方法
海水鱼白点病治疗方法 综述
综述